This post was inspired by an MD using PEMF who was convinced that PEMF was enabling saltatory conductance by myelinated fivers. In the original experiments of Luigi Galvani and his frog legs in which nerves were impaled with electrodes ]1] This post is written for MDs and others with a very good background in medical physiology, chemistry, and biophysics. Background information is provided for the well motivated lay person. The reader may end up questioning what they thought they knew about nodes of Ranvier and conduction of impulses along myelinated axons. My conclusion in this thought and peer reviewed literature journey is that that the MD was insightful but mistaken. Their intuition was correct about cytosolic and sub myelin “two cable” conductance. Mathematical modeling suggests that capacitors exist in both fractions as well. The PEMF Faraday Law of Induction geometry is just wrong. This post proposes that PEMF may reset the axon circuit.
Saltatory conductance at Nodes of Ranvier cartoon.
The Node of Ranier Panel A image came from Wikimedia Commons originally appearing in Min Y., Kristiansen K., Boggs J. M. at al (2009). “Interaction forces and adhesion of supported myelin lipid bilayers modulated by myelin basic protein”. Proceedings of the National Academy of Sciences 106 (9): 3154–3159.

Panel B I find the concept of saltatory (leap frog) conductance and the way it is explained in the medical school physiology course woefully inadequate. The anions and cations on the outside of the cell membrane as well as inside the cell do not enter the discussion as to how one node knows what the upstream node is doing. One of the exercises in graduate school was to calculate the number of positive chargers per area of a node due to the action of the Sodium Pump that pumps out three Na+ out for every 2 K+ pumped in. Because of electro-neutrality rules these Na+ stay in the vicinity. All positive charges must be balanced by negative charges. Outside the cell the negative charges tend to be chloride anions. Inside the cell, protein amino acid side changes are a source of negative charges. Some of the blue circles inside the cell should be connected to symbolize a proteins. Furthermore, outside the cell there are usually negatively charged glycated (sugar polymers) proteins that provide a gel for cations to concentrate from the bulk extracellular solution. Panel C My hypothetical model follows the same principal as Newton’s pendulum that has been morphed into another Wikimedia Commons image. Little did I know… when I started this thought journey, the following aligns with a ground breaking paper that suggests a layer of charges beneath the myelin that act as a double cable [2].
Here is a copper wire. The electrons are delocalized about the nuclei and flow to one end when a positive voltage drop is applied.
No Wikipedia links to mathematics and philosophy intensity content written by advanced academics. Instead just some images. A copper wire coming out of a blue insulating cover. The red balls represent the nuclei of copper atoms. Valence electrons circulate freely among the nuclei. When a positive charge is applied to one end the electrons slosh towards this charge. The bigger the charge (voltage) drop, the more electron sloshing , bumping into one another, towards the anode, positive end. Some readers may remember the equation V = I R from high school physics. I is the current and R the resistance. If the copper nuclei hold tightly to the outer orbit electrons, the “sloshing” of electrons is not so free. The wire has resistance.

What happens when charged particles are just floating around a charged surface much like electrons are floating about copper atom nuclei in a copper wire?

Imagine internodal myelinated regions coated with negatively charged glycoproteins. Could these ions respond to PEMF via Faraday’s Law of induction?
Zeta potential is the electrical potential at the slipping plane, the region at which the counter ions associated with a charged surfaced start to resemble those of the bulk solution. The Stern double layer is a layer of charged particles associated with a surface of opposite charge. After this is a more sparsely populated layer of particles with the same charge as the surface. The first layer shields the second of repulsive forces of like charge.
A Na+ at the node thought experiment
- We are taught that extracellular fluid is 150 mM Na+.
- Using Avogadro’s number of 6.02214076×1023 mol−1 we can calculate 0.9×1023 Na+ in a 10×10 cm square of fluid. cm is also 1 liter.
- each time we decrease the length of the square 10x we decrease the number of Na+ 1000x because 10 x 10 x10 =1000
- If we go down to 10-9 m or nm squares we have a reduction factor of 3x 7 = 21
- subtract exponents: 23-21=2
- 0.9×102 Na+ for 90 Na+ per cubic nm.
- Mähler & Persson 2011 give the hydrated radii of six water molecule coordinated Na+ and K+ as 1.07 and 1.38 Å respectively. There are 10 Å in 1 nm. There are 1000 square in our cubic nm. Only 90 of these contain Na+!
Colorado State has some assembled a list of conductivity of common salts as a function of concentration. Conductivity is the opposite of resistance. As concentration increases so does conductivity. If the body can concentrate ions above 150 mM…
the double cable model [2]
historical prelude and what a capacitor does
Physiopedia saltatory capacitance by Dr. Ichiji Tasaki. This post offers a nice historical background of how scientists came to view that myelin acts as a capacitor. A capacitor stores electrical energy by accumulating electric charges on two closely spaced conductive surfaces that are insulated from each other by a dielectric medium. Ceramics and mica are commonly used dielectrics. Wikipedia authors provide a hydraulics analogy that has a close cousin of Windkessel blood vessels in our own bodies. The aorta and other major blood vessels are highly elastic and expand as blood flows through them with each heartbeat. They release that stored blood during diastole. Flow through the aorta and through a capacity has an out of phase with our heartbeat and an AC voltage source, respectively. .Here is a link to an image of how “current” through a capacitor is out of phase with an AC voltage source.
Along with capacitors , there are two cables/conductors in the axon! [2]
This study [2] challenged the traditional notion derived from work with frog sciatic nerves that myelin fibers are insulated single cable networks. Other ideas have been proposed that two cables may exist: one running inside the myelin and the other running just underneath it. [6] Using a combination of patch clamping and optical techniques it was concluded that a double cable model best fits the data. Myelin remained the insulator with a conductive layer of ions just beneath it. Fig 2 offers a nice summary of what this paper was all about. Panel A describes the patch clamping along the length of a myelinated axon. Panel B is the traditional single cable model. Panel C is the double cable model in which a resistor, really a conductor, is underneath the myelin capacitor. This paper has some very nice electron micrographs that supports the notion that such a space exists for ions to move about. Much of this paper was a round about of fitting models to data collected with patch clamp electrodes and potential sensitive dyes. It should be noted that the data refined equivalent circuit mathematical model allowed the cytosol also acting as a capacitor.
the capacitor is in the axon cytosol ?[3]
Tsubo & Kurokawa 2018 conducted an in slico study testing the hypothesis that the dielectric constant of axon fluid functions as a capacitor in addition to the one of the myelin sheath. A Wikipedia of dielectric constant redirects to relative permittivity two capacitors in which the stuff between the two plates of the reference capacitor is a vacuum. The test material has a frequency dependency. If we are to trust Wikipedia authors, at 25oC and 1MHz ethanol, methanol, and water have diectric constants of 24.3, 32.7, and 78.4. All three very small molecules have “OH” groups and an uneven distribution of partial charges that can orientate in and electric field. The smaller the molecule, the easier it is to spin around. What if the AC source is the action potential of the neuron? Do the “much bigger than water” proteins have time to respond?
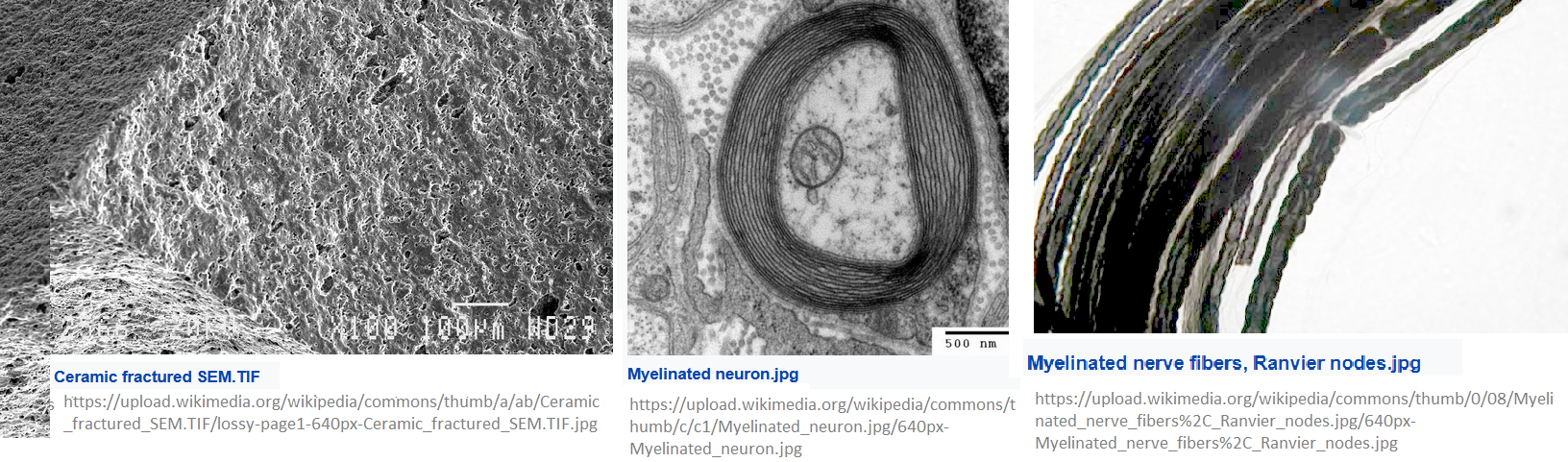
The cytosol, shock full of proteins with flexible bonds and charges. The featured image compares electron microscope images of a ceramic that is used in electronic capacitors with two Wikipedia Commons images of myelin and axon fluid. The myelin looks more like a capacitor material with nooks and crannies to load charged particles. Skipping some mathematically intense theoretical modeling, here is the conclusion:
The wavelength of an action potential is 0.885 m at a frequency of 2 kHz, and the action potential is affected by an axonal transmission characteristic to approach the same level of length of the motor and sensory nerve. However, the properties of the myelin sheath make it a good transmission line, as mentioned in section 1.4. The morphology of the myelin sheath is a good complement to the extremely short wavelength of the action potential of the carrier of axon fluid. On the other hand, it becomes the domain where impedance is very low, where the membrane is very thin because the node of Ranvier does not have a sheath. However, the length of node of Ranvier region (2 μm) is very short compared with the wavelength (0.885 m), and it is less than 1/440000. Therefore, we thought that the quantity of energy loss of one wavelength of action potential by node of Ranvier region is negligible. Rather, it is thought that this very short length is useful to ensure a low capacitance Cn of the node of Ranvier. “
Multiple Wikipedia authors have compiled a page on the action potential that give the length of just one action potential in the msec time range of an action potential itself. The temporal spacing of action potentials is far more difficult to fact check because the truth is all over the place depending on the system in question.
Kirkcaldie & Collins the crowded gel of “axonal fluid” [4]
Tsubo & Kurokawa 2018 [2] was a very nice thought provoking study. For those that have worked with isolated cytoskeletal proteins vary from gels to very tough fiber. The nature of the Kirkcaldie & Collins 2016 review [4] came from the perspective of why don’t peripheral nerves get injured more in the stretching, compressing, and out right blunt force trauma? The elasticity of this gel was the take home theme.
- Spectrins (think red blood cells) form a cortex with actin (in red blood cells too) just inside the axolemma, Micro filament actin protects the cell membrane of the axon. Actin is subject to cross-linking and,along with spectrin, anchorage to the axon membrane.
- Neurofilament’s primary function is to protect the brittle macrotubules. NF (light, medium, and heavy) have side arms to structure the axoplasm into a gel. .
- Microtubules, the larger class of filaments, are conduits for synaptic vesicles containing neurotransmitters and mitochondria.
The authors have an excellent cartoon for conceptualization of what makes our axons so strong in health. The “take home” message for this post is that these filaments are crowded, strong, and likely to be covered with counter ions that can theoretically response to action potentials and PEMF.
Myelin sheath structure [5]
This combination review and nerve injury study focuses on the importance of myelin basic protein in the formation of the myelin sheath. The C-fiber Remak bundle bridges the gap between capacitance and PEMF improvement of nerve conductance and nerve injury repair. Wikipedia authors claim that the Remak Schwann cells are electrically chemically coupled to the action potentials. Human Physiology Academy Remak Bundle shows a normal Schwann myelinated axon with about 10 myelin sheaths. Next to it is a Schwann cell engulfing five neurons..
UniProt, exploring the potential dipole nature of MBP
This is the UniProt sequence for human myelin basic protein
- Length304
- Mass (Da)33,117… There are four isoforms with molecular weights of 21.5, 18.5, 17, and 14 kDa.

Liu 2019 [6] introduction to MBP
- MBP undergoes acetylation, methylation, and phosphorylation during the formation of a 3D structure [6]
- Fig 1 shows positive charge clusters on MBP and negative charges in the glial cell membrane. These electrostatic interactions are the driver of myelination.
- Myelin layers are 20-30% protein and 70-80% lipid.
- In the double-bilayer structure, the 2 layers of cell membranes are separated by 3 to 8 nm and are filled by the cytoplasm of glial cells.
- Fig 2 is composed of transmission electron micrographs of the optic nerve, oculomotor nerve, and sciatic nerve
Repair of a spliced together sciatic nerve
The narration of Liu 2019 [6] then turned to peripheral nerve injury repair. This could be of interest to those using PEMF for nerve injury repair. They used End-to-end anastomosis of the broken sciatic nerve was performed in a rat model of nerve injury (10 mm), An anastomosis is the surgical connection of separate or severed tubular hollow tissues to form a continuous channel, e.g. the intestine, blood vessels, and the sciatic nerve..
- Fig 3 shows marker proteins in the distal and proximal end of the growth cone. Nerve growth factor receptor (NGFR) and MBP are two of four proteins examined. NGFR was abundantly expressed in the distal region of the growth cone.
- Fig 4 TEM images 30 days after surgery. Some abnormal sheath condensation.
- Fig 5 The expression levels of lipophilin, MBP, GFAP, NF200, and NGFR P75 relative to normal sciatic nerve in the end-to-end anastomosis, autologous nerve transplantation, and nerve tube transplantation groups and in primary cultured Schwann cells
The discussion then turned to how injured nerves communicate with Schwann cells. The expression of MBP is tied to this process.
Postlude communication with a retired expert
Dr George Harauz spent spent much of his career studying the structure of MBP and its interactions with phospholipid membranes. His thought was that MBP has a hairpin or paperclip structure. A dipole moment in the membrane seems to be unlikely due to all of the negative charges of the phospholipids and positive charges on MBP. The game changing study of Gohen 2020 [2] assumed that Na+ was the conducing ion. Na+ being the counter ion of negatively charged phospholipids makes perfect sense. One would hypothesize that Cl– would be a reasonable counter ion for the lysines and arginines of BBP.

A rudimentary model based on the work of Cohen 2020 [2] proposing Na+ beneath the myelin sheaf. The new concept is that there are both negatively charged Cl– ions (red circles) and Na+ ions (blue circles). Because the membrane spaces have negative (phospholipids) and positive charges (MBP) we can speculate that there cations and anions that can move in an electrical potential difference such as an action potential. Kirkcaldie & Collins 2016 [4], and Liu 2019 [5] describe the importance of MBP in this region. Not in this very crude cartoon is the possibility that proteins in this peri-axonal space may also orientate in an electrical field as proposed by Tsubo 2018 for axonal fluid. [3]

Do these perpendicular electrical fields start out that way and then curve around taking the path of least resistance? Dennis Kunkel (click to open in new tab) has a good longitudinal scanning electron micrograph of a myelinated axon. Imagine the magnetic field going into the screen. Imagine electric fields going up and down the long axis of the axon. Imagine an electric field going in and out of layers of myelin sheaths? Can they even do this when they hit the acyl chains of the lipid bilayer?
References
- Piccolino M. Luigi Galvani and animal electricity: two centuries after the foundation of electrophysiology. Trends Neurosci. 1997 Oct;20(10):443-8. doi: 10.1016/s0166-2236(97)01101-6. Erratum in: Trends Neurosci 1997 Dec;20(12):577. Sci-Hub free paper
- Cohen CCH, Popovic MA, Klooster J, Weil MT, Möbius W, Nave KA, Kole MHP. Saltatory Conduction along Myelinated Axons Involves a Periaxonal Nanocircuit. Cell. 2020 Jan 23;180(2):311-322.e15. PMC free paper
- Tsubo T, Kurokawa M. Verification of the effect of the axon fluid as a highly dielectric medium in the high-speed conduction of action potentials using a novel axon equivalent circuit. Biophys Physicobiol. 2018 Oct 25;15:214-228. PMC free paper
- Kirkcaldie MT, Collins JM. The axon as a physical structure in health and acute trauma. J Chem Neuroanat. 2016 Oct;76(Pt A):9-18. Sci-Hub free paper
- Liu B, Xin W, Tan JR, Zhu RP, Li T, Wang D, Kan SS, Xiong DK, Li HH, Zhang MM, Sun HH, Wagstaff W, Zhou C, Wang ZJ, Zhang YG, He TC. Myelin sheath structure and regeneration in peripheral nerve injury repair. Proc Natl Acad Sci U S A. 2019 Oct 29;116(44):22347-22352.. PMC free paper
Leave a Reply